SAMHSA’s six key principles for a trauma-informed approach to Trauma
SAMHSA’s six key principles for a trauma-informed approach to Trauma
SAMHSA’s six key principles for a trauma-informed approach to Trauma
Instructions:
Students will choose a specific type or aspect of trauma that interests them.
Using scholarly references, the paper will include a comprehensive overview of the type (or aspect) of trauma, a critical analysis of the biopsychosocial factors affecting trauma exposure and recovery, a brief discussion of the best practices for intervention, application of SAMHSA’s six key principles for a trauma-informed approach, and personal reflections.
Write the final paper using the following headings to structure your writing, and include references page.
SAMHSA’s six key principles for a trauma-informed approach to Trauma
Introduction – Briefly describe the focus and purpose of the paper. The introduction should give your reader an understanding of the dimension of trauma you will focus on, and the level of social work intervention you will address (1/2 page).
Overview of Trauma – Drawing from the professional literature, provide a detailed examination of the type and nature of the trauma, prevalence, specific characteristics, its impact across several domains (psychological, emotional, spiritual, etc.).
Describe any relevant DSM-5 or mental health diagnoses for a person exposed to this type of trauma, if appropriate (2-3 pages).
Biopsychosocial Factors- Discuss how the context of a person’s environment influences an individual or community’s exposure and response to this type of trauma.
Evaluate both risk and protective factors impacting one’s exposure to and response to trauma.
Include personal characteristics, as well as a discussion of any social, cultural and economic justice factors that may affect the client’s vulnerability to this trauma, the availability of and access to services, and any unique challenges in recovery (e.g., social class, gender, ethnicity, age, religion, etc.) (1 ½-2 pages).
Intervention/Core Trauma Competencies – Briefly review the professional literature on best practices for intervening with this type of trauma, whether micro and macro responses (1 page).
SAMHSA’s six key principles for a trauma-informed approach to Trauma
Discuss how a practitioner would apply SAMHSA’s six key principles in treating this type of trauma (safety, trustworthiness, collaboration, empowerment, peer support, culture, gender & historical issues). Be specific when describing the application of these principles in practice (2- 3 pages total).
Student should include a minimum of six (6) references from professional literature regarding best practices, including a minimum of three (3) peer-reviewed journals.
To get a custom answer click order now and apply the 25% discount voucher: MBW25
Ivy, age 23, is a white woman who graduated from college last year. She began working as an accountant one month after graduating. Approximately two months ago, she moved into a two -bedroom apartment with another woman who works at the same accounting firm.
She states that her roommate recommended that she sees a doctor to find out if she has anemia or “some sort of fatigue syndrome.” She states that she has felt “restless” and “on edge” for most of the past nine months.
Case Study Questions – Anxiety Disorder
She becomes easily fatigued and irritable and has difficulty concentrating and falling asleep. She states that sometimes her mind “just goes blank,” and she is worried that her work performance is no longer excellent.
Although she has been “a worrier from the day I was born,” now she worries more than she ever has and feels nervous “all the time.” Ivy reports that she has a good relationship with her boyfriend, but they do not get to see each other very often because he is attending graduate school 100 miles away.
She reports having a satisfying sexual relationship with him. She denies having any problems with relationships with her parents, roommate, or peers. She denies having any financial worries unless she is fired from her job for poor work performance.
Case Study Questions – Anxiety Disorder
She reports that she has always been healthy and has taken good care of herself. The only medication she takes is birth control pills, which she has taken for the past four years without any adverse effects.
I started some already but I need help. Read the case study on generalized anxiety disorder (GAD) and answer the8 questions related to reading and other outside sources. I would like to try to keep what I have and my sources and just build from there. Do you think that will be OK?
Generalized Anxiety Disorder (GAD) is one of the mental health disorders set apart by a feeling of worry, fear, restlessness, or feeling of impending doom. According to the World Health Organization (WHO), 1 in 13 persons suffers from anxiety.
Case Study Questions – Anxiety Disorder
The WHO reports that anxiety disorders are the most common mental disorders worldwide with specific phobia, major depressive disorder, and social phobia being the most common anxiety disorders (WHO, 2021 ? find here >> https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2174588/ ).
Anxiety can usually be self- diagnosable as with Ivy in this case study. Women, unlike men, are twice as likely to be affected with GAD (Woo & Robinson, 2020).
Just making it through a simple day causes anxiety and like with Ivy, the 6.8 million adults with GAD do not know how to stop the worry cycle and feel out of control or even things are beyond their control.
GAD can interfere with a person’s daily events and actives as with Ivy. For nine months Ivy has had trouble concentrating at work and now worries about her job security and performance (ADAA, 2021).
SSRIs can be used as the first-line medication for Ivy’s anxiety disorder. SSRIs cause more serotonin- 5-HT- in your brain to be available therefore improving anxiety as well as mood (Woo & Robinson, 2020).
Question 1. List specific treatment goals for Ivy. Treatment goals for Ivy include staying compliant with her new medication regime of SSRI- Lexapro 10 mg PO once daily, keeping follow-up appointments to assess and evaluate new medication responses and therapeutic outcomes, better sleep, increase quality in work performance.
Participate in psychotherapy (cognitive-behavioral/solution-focused) **For #1 need to include National guidelines for treatment plans and options- please look at US Preventive Services Task Force https://www.uspreventiveservicestaskforce.org/uspstf/recommendation-topics
American Psychiatric Association Practice Guidelines
https://psychiatryonline.org › guidelines
American Psychiatric Association (APA) practice guidelines provide evidence-based recommendations for the assessment and treatment of psychiatric disorders.
Case Study Questions – Anxiety Disorder
Question 2. What drug therapy would you prescribe? Why? As mentioned before, SSRIs are used as the first-line medication with a new diagnosis of GAD with a safer and lower side-effect profile (Woo & Robinson, 2020).
I would start Ivy on Escitalopram (Lexapro) 10mg PO once daily, this dose may be increased after response, and tolerability is assessed to a maximum of 20mg once daily (Lexicomp, 2021).
Lexapro chemical action blocks the transport mechanism that helps return 5-HT that is unbound from returning into the presynaptic neuron therefore more 5-HT is increased and available for the brain to blind to the postsynaptic serotonin receptor.
This in turn can decrease symptoms of anxiety (Woo & Robinson, 2020). The effectiveness of the different SSRIs over each other measures the same with similar outcomes (Woo & Robinson, 2020) (Riley et al., 2009).
Question 3. What are the parameters for monitoring the success of the therapy?
Question 4. Describe specific patient monitoring based on the prescribed therapy.
Question 5. List one or two adverse reactions for the selected agent that would cause you to change therapy. Look and read UpToDate and Lexicomp under Escitalopram (Lexapro)
Question 6. What would be the choice for second-line therapy? According to Lexicomp Zoloft would be the second-line medication for Ivy.
Question 7. What dietary and lifestyle changes should be recommended for this patient? According to Woo & Robinson (2020) exercise (no matter the person’s intensity) can be a constrictive part in reducing the symptoms of anxiety.
Exercises such as brisk walks, yoga, and meditation can be helpful for Ivy’s treatment. Cognitive-behavioral therapy (CBT) can also be beneficial to the person with GAD as well as Ivy’s boyfriend or even her roommate.
Case Study Questions – Anxiety Disorder
CBT uses psych-social interventions aimed to improve anxiety by drawing focuses on changing cognitive distortion such as thoughts, beliefs, and attitudes as well as managing coping strategies that target personal problem solving and enhancing overall wellbeing (Menzies, et al., 2016).
Alcohol consumption should be avoided for Ivy. Alcohol can enhance the side effects causing increased psychomotor impairment[UptoDate,2021].
Question 8. Describe one or two drug-drug or drug-food interactions for the selected agent.
To get a custom answer click order now and apply the 25% discount voucher: MBW25
Identify the target symptoms for a child or adolescent from your practicum site. Must be a different case then the first one.
2 State the DSM-5 diagnosis with evidence.
Identify two differential diagnosis with rationale.
Interventions
List non-pharmacological interventions with rationale.
Psycho-pharmacological intervention with rationale.
Include a sample prescription
Provide a link to the medication education sheet to be given to the family/patient.
Provide a link to the appropriate diagnostic sheet to be given to the family/patient.
List additional assessments required ie screening tools, labs with rationale.
Name a community resource / referrals for the patient/family. (It can be a web site for a support group; literature, organizations, educational support, or a list of community resources etc.)
Korean Students and Mental Health to Parents Effects of Strategy Type to Disclose Korean Students and Mental Health to Parents
Korean Students and Mental Health to Parents
Research Question: (please rewrite in a more advanced way if possible)
Imagine you are a student with mental health issues. Your psychiatrist has deemed that you have depression, bipolar disorder, or any other type of disorder and may need to take medication. How would you tell your parents? Would you tell them at all?
Participants: 15 born and raised Korean students will be given a real-life situation in which they must Need to reference:
Formula: Wx = D(S, H) + P(H, S) + Rx
Wx is the weight which refers to the weightiness of the act.
D(S, H) symbolizes the distance between the distance and speaker
P(H, S) + R stands for the power o the hearer over the speaker
Rx implies the degree of the imposition of the act
Needs to include:
A section on Korean culture regarding mental health Quantitative Measurement
We can write this or a similar paper for you! Simply fill the order form!
Quantitative Molecular Imaging of Neurotransmitters Direct Targeted Quantitative Molecular Imaging of Neurotransmitters in Brain Tissue Sections
Quantitative Molecular Imaging of Neurotransmitters
Mohammadreza Shariatgorji,1 Anna Nilsson,1 Richard J.A. Goodwin,1,2 Patrik Ka® back,1 Nicoletta Schintu,3
Xiaoqun Zhang,3 Alan R. Crossman,4 Erwan Bezard,5 Per Svenningsson,3 and Per E. Andren1,
* 1Biomolecular Imaging and Proteomics, National Center for Mass Spectrometry Imaging, Department of Pharmaceutical Biosciences,
Uppsala University, P.O. Box 591 BMC, 75124 Uppsala, Sweden
2AstraZeneca R&D, Alderley Park, Macclesfield, Cheshire SK10 4TF, UK
3Center for Molecular Medicine, Department of Neurology and Clinical Neuroscience, Karolinska Institutet and Karolinska University Hospital, 17176 Stockholm, Sweden
4Faculty of Life Sciences, University of Manchester, Manchester M13 9PL, UK
5Universite¥ de Bordeaux, Institut des Maladies Neurode¥ ge¥ ne¥ratives, UMR 5293 Bordeaux, France
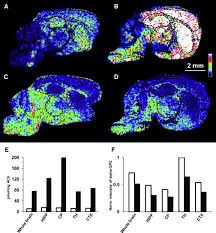
Current neuroimaging techniques have very limited abilities to directly identify and quantify neurotransmitters from brain sections. We have developed a molecular-specific approach for the simultaneous imaging and quantitation of multiple neurotransmitters, precursors, and metabolites, such as tyrosine, tryptamine, tyramine, phenethylamine, dopamine, 3-methoxytyramine, serotonin, GABA, glutamate, acetylcholine, and L-alpha-glycerylphosphorylcholine, in histological tissue sections at high spatial resolutions. The method is employed to directly measure changes in the absolute and relative levels of neurotransmitters in specific brain structures in animal disease models and in response to drug treatments, demonstrating the power of mass spectrometry imaging in neuroscience.
Quantitative Molecular Imaging of Neurotransmitters Introduction
Small-molecule neurotransmitters such as the catecholamine dopamine (DA), the amino acids g-aminobutyric acid (GABA) and glutamate (Glu), and acetylcholine (ACh) are critical chemical messengers that transmit signals between neurons. Changes in their concentrations are associated with numerous normal neuronal processes, such as sleep and aging, but also several disease states, including Alzheimerís disease, Parkinsonís disease (PD), depression, drug addiction, and attention deficit hyperactivity disorder. A better understanding of their relative abundance and distribution would provide insights into these complex neurological processes and disorders. At present, researchers rely on indirect histochemical, immunohistochemical (IHC), and ligand-based assays to detect these small-molecule transmitter substances (de Jong et al., 2005; Falck et al., 1962;
Jones and Beaudet, 1987). The antibodies used in IHC often have major limitations, such as an inability to distinguish between different transmitters (Keenan and Koopowitz, 1981).
Nuclear medicine imaging is a widely used tool for indirect visualization of the distribution, abundance, and activity of neurotransmitters in the brain, relying on radiolabeled tracers that emit gamma rays, which are detected using positron emission tomography (PET) or single photon emission computed tomography scanners (Pimlott and Sutherland, 2011). Developing appropriate labeled compounds is often complicated, particularly for studying endogenous chemical messengers (Badgaiyan, 2011). There is often an inability to discriminate between the labeled parent compound and a metabolite retaining the label. Imaging techniques that can directly visualize the localization of neurotransmitters and simultaneously quantitate them without relying on a label would thus represent a scientific breakthrough. Another significant advance in the molecular analysis of neurological tissues would be the development of a multiplex method that enables the simultaneous detection of multiple endogenous compounds and pharmaceutical agents.
MALDI mass spectrometry (MS) imaging has been used for direct molecule-specific compound detection, distribution mapping, and identifying molecular species in tissue sections (Caprioli et al., 1997; Cornett et al., 2007). It can generate multiplex full mass spectrum data at near-cellular spatial resolution.
Crucially, it provides a powerful tool for in situ visualization of target compounds and for determining their abundance and spatial distribution without requiring a great deal of a priori information.
Moreover, the data gathered during such experiments can be used to simultaneously identify unknown endogenous molecular changes due to disease or degeneration. MALDI-MS can be used for the direct analysis of targets, ranging from small molecules to large proteins (Schwamborn and Caprioli, 2010).
However, despite some notable successes, it is relatively insensitive toward certain compound classes and sometimes suffers from signal interference caused by matrix clusters/fragments (Sugiura et al., 2012; Ye et al., 2013). To date, it has found only limited applications in the study of neurotransmitters due to their poor ionization efficiencies. As such, the development of a reagent that would enable selective neurotransmitter derivatization while also facilitating their detection by MS and acting as an efficient matrix to assist their laser-induced desorption and ionization would be a significant breakthrough. Neuron 84, 697ñ707, November 19, 2014 ™2014 Elsevier Inc. 697
Here we describe a unique approach for the direct and absolute quantitative imaging of classical and common neurotransmitters in histological brain tissue sections. We report an example of the simultaneous mapping of biogenic amines and amino acid chemical messengers following in situ derivatization in brain tissue sections. The method is employed to simultaneously measure changes in the levels of neurotransmitters, including tyrosine, tryptamine, tyramine, phenethylamine, DA, 3-methoxytyramine (3-MT), serotonin (5-HT), GABA, and Glu, in specific brain structures of primates and rodents with and without DA depletion and L-3,4-dihydroxyphenylalanine (L-DOPA) treatment. In addition, we have developed a method that can be used to image and quantitate ACh and an ACh precursor, L-alpha-glycerylphosphoryl-choline (alpha-GPC) in brain sections, as well as to study activity within cholinergic pathways by measuring changes in ACh concentrations, imaged at a 15 mm spatial resolution. Together, these experiments demonstrate a significant advancement in the molecular imaging arsenal available to neuroscientists.
We can write this or a similar paper for you! Simply fill the order form!
The domain of social work in mental health is that social context and social consequences….discuss.
Your assignment must follow these formatting requirements:
Be typed, double spaced, using Times New Roman font (size 12), with one-inch margins on all sides; citations and references must follow APA or school-specific format. Check with your professor for any additional instructions.
Include a cover page containing the title of the assignment, the student’s name, the professor’s name, the course title, and the date. The cover page and the reference page are not included in the required assignment page length.
We can write this or a similar paper for you! Simply fill the order form!
The Case Study is: SUBSTANCE USE DISORDER (Case study 4)
(1) Explain what the purpose of a clinical assessment is.
(2) Identify a commonly used instruments clinicians or psychologists use to screen for a particular disorder and why it would be used. Note, the DSM-5 is not a screening instrument. An instrument helps to identify if a patient is experiencing the symptoms listed in the DSM-5. An example of an instrument is the Beck Depression Inventory that screens for symptoms Major Depressive Disorder, or the PC-PTSD that is designed to screen for post- traumatic stress disorder.
(3) During a clinical assessment, why would a clinician consider the level of severity of a patient’s symptoms?.
We can write this or a similar paper for you! Simply fill the order form!
how does the media represent/view offenders (pick one type of crime) with mental health disorders or those who consume illegal substances”
SUBJECT: Criminology, ”how does the media represent/view offenders (pick one type of crime) with mental health disorders or those who consume illegal substances” (this is a working title and i do not have a problem with it being tweaked as long as it is of a criminology nature)
Your assignment must follow these formatting requirements:
Be typed, double spaced, using Times New Roman font (size 12), with one-inch margins on all sides; citations and references must follow APA or school-specific format. Check with your professor for any additional instructions.
Include a cover page containing the title of the assignment, the student’s name, the professor’s name, the course title, and the date. The cover page and the reference page are not included in the required assignment page length.
We can write this or a similar paper for you! Simply fill the order form!
This Abnormal Behavior paper is needed for my Mental Health in Criminal Justice class.
Write a 700- to 1,050-word paper in which you examine the field of abnormal psychology. Address the following items:
• Describe challenges related to classifying normal versus abnormal behavior.
• Analyze the psychosocial, biological/medical, and sociocultural theoretical models related to the development of abnormal behavior.
• Describe the relationship between abnormal behavior and criminal behavior.
• Explain how correctional institutions have been affected by abnormal behavior.
We can write this or a similar paper for you! Simply fill the order form!




:max_bytes(150000):strip_icc()/gad-causes-risk-factors-1392982-488cca43a0484ff7804288f6129035f9.png)